
İskelet kaslarının kasılma yapısı ve mekanizması. Hayatımızdaki mineraller (bunu bilmek güzel) İskelet kaslarının çalışması için iyonlara ihtiyacı vardır.
İskelet kasını oluşturan uzun ince kas lifleri, çok sayıda hücrenin birleşmesi ile ontogenez sırasında oluşan dev hücrelerdir. Bir yetişkinde 5 cm uzunluğa ulaşabilirler! Böyle bir hücrede çok sayıda çekirdek, doğrudan sitoplazmik zarın altında bulunur ve sitoplazmanın ana kısmı, karakteristik bir enine çizgi ile tüm hücre boyunca (1-2 um kalınlığında) uzayan miyofibrillerden oluşur (Şekil 3). Miyofibrilin bu “renklendirmesi”, her biri iki paralel, kısmen örtüşen filament içeren sarkomerler tarafından verilir: koyu bir şerit oluşturan ve bir kenardan diğerine uzanan kalın miyozin filamentleri ve ince aktin filamentleri. açık şerit bölgesinde ve kısmen koyu şeritler alanına uzanır (Şekil 4). Bir sarkomer diğerinden bir Z diski ile ayrılır.
Pirinç. 3. İskelet kası hücresinin küçük bir bölümünün şeması (kas lifi)
Pirinç. 4. A. Bir tavşan iskelet kası hücresinin (düşük büyütmede) boyuna kesitinin elektron mikrografı. Düzenli enine çizgilenme görülebilir. Hücre birçok paralel miyofibril içerir (bkz. Şekil 3). B. Aynı fotoğrafın iki bitişik miyofibrilin segmentlerini ve sarkomerin ayrıntılarını gösteren küçük bir bölümü. B. Elektron mikrografında görülen koyu ve açık bantların kökenini açıklayan tek bir sarkomerin yapısal diyagramı
Sitoplazma, miyofibrillere ATP şeklinde enerji sağlar - aktif olarak çalışan bir kasta birçok mitokondri bulunur. Ek olarak sitoplazma glikojen, fosfokreatin, glikolitik enzimler içerir.
İskelet kası, ATP'de depolanan kimyasal enerjiyi çok yüksek verimle mekanik enerjiye dönüştürür - ısı olarak yalnızca %30-50'si kaybolur. (Karşılaştırma yapmak gerekirse, bir araba motoru benzin yakarken tipik olarak %80-90 ısı kaybeder.)
Kas kasılması sırasında, miyozin tarafından aktin bağlanması nedeniyle kalın ve ince filamentler birbirine göre kayar (Şekil 5).
Pirinç. 5. Kayma iplikleri ilkesine göre kas kasılma sürecini gösteren şema: kalın ve ince filamentler kendi uzunluklarını değiştirmeden birbiri üzerinde kayar
Kalsiyum iyonları, kas kasılmasının fizyolojik düzenleyicisi olarak hizmet eder. Dinlenirken, kalsiyum iyonlarının aktif taşıma sistemi çalışır ve bir sinir impulsunun etkisi altında serbest bırakıldıkları ve kas kasılmasını sağlayan bir tür depolamada birikir.
Kalsiyum iyonu taşıma sistemi, ATP'nin enerjisiyle çalışır. Kasta bulunan ATP miktarı, kasılma aparatının çalışmasını saniyenin çok küçük bir bölümünde sürdürmek için yeterlidir. Bir kas nasıl daha uzun süre çalışır? Kasın, enerjiyi evrensel ATP'den daha yüksek enerjili fosfat grupları taşıyabilen fosfokreatin veya kreatin fosfat şeklinde depoladığı ortaya çıktı. Fosfokreatin ATP'yi geri yükler, böylece kas kasılması için bir enerji akışı sağlar. Ancak çalışan bir kasta fosfokreatin rezervleri hızla tükenir ve bu da ATP içeriğini azaltır.
Daha uzun bir fiziksel yük ile, kaslara glikoliz nedeniyle enerji sağlanır - enzimlerin etkisi altında karbonhidratların ATP şeklinde enerji birikimi ile parçalanması. Kastaki kreatin depoları tükendiğinde, kas kasılmasının enerji yükü azalır, bu da çalışan kasta glikoliz, trikarboksilik asit döngüsü ve oksidatif fosforilasyonun uyarılmasına yol açar.
Oksijen yokluğunda, bir karbonhidrat molekülü parçalandığında, iki molekül laktik asit (veya laktat) ve iki molekül ATP oluşur. Bununla birlikte, glikoliz için kas glikojeni kullanılırsa, iki laktat molekülü ve üç ATP molekülü üretilir.
Glikojenin kaslarda ve karaciğerde depolanan ana rezerv polisakkarit olduğu bilinmektedir. Kaslarda ve karaciğerde düşük glikojen seviyeleri ve kanda serbest glikoz varlığı ile glikojen sentezi için kullanılır. Tersine, vücut glikoliz işlemleri için bir enerji kaynağına ihtiyaç duyduğunda, glikojen başarıyla kullanılır.
Kas sistemi diğer vücut sistemlerine göre en gelişmiş olanıdır. Kasların çalışmasını sağlamak için, bir kişinin kreatin fosfattan (fosfokreatin), glikojen ve glikoz ve yağ şeklindeki karbonhidratlardan alabileceği büyük miktarda enerjiye ihtiyaç vardır. Bu üç tür enerji taşıyıcısı, kullanımları sırasında açığa çıkan enerji miktarı ve her birinin ne kadar süre "yakıt" kaynağı olarak hizmet edebileceği açısından birbirinden farklıdır. Kaslara enerji sağlayan mekanizma bu şekilde düzenlenir (Tablo 2, 3). Uzun süreli yoğun olmayan çalışma sırasında, oksidatif işlemler sırasında yağların veya karbonhidratların kullanıldığı ve biraz daha yüksek yoğunluklu çalışma sırasında anaerobik glikoliz mekanizmalarının kullanıldığı iyi bilinmektedir. Çok yoğun bir kısa süreli yük ile kas çalışması fosfojenler tarafından sağlanır. Buna göre enerji kaynaklarının her birinin kendi enerji maliyeti vardır ve belirli koşullar altında kullanılmaktadır (Tablo 4).
Diğer organlarla karşılaştırıldığında, çalışan kaslar karbonhidratların aerobik oksidasyonu için çok büyük miktarda oksijen tüketir (Tablo 5).
Tablo 2. 70 kg ağırlığındaki insan vücudundaki enerji rezervleri
Tablo 3. Çeşitli substratları ve katabolizma yollarını kullanan insan iskelet kaslarının maksimum olası gücü
Tablo 4. Egzersiz sırasında çeşitli kaynaklardan maksimum enerji üretim hızı
Enerji kaynağı |
Maksimum kas hızı |
Enerjiden zengin fosfat bağlarının oluşumunun kastaki miktarı, mmol/kg, mmol/sn/kg |
Maksimum üretim hızı, kcal/h/kg |
Maksimum hızı koruma zamanı |
Kreatin fosfat |
||||
anaerobik glikoliz |
||||
Glikoz ve glikojenin aerobik oksidasyonu |
||||
aerobik oksidasyon |
Sınırsız |
Sınırsız |
Tablo 5. Dinlenme ve ağır çalışma sırasında çeşitli insan organlarının nispi oksijen tüketimi *
İskelet kası tonu. Dinlenirken, iş dışında, kaslar asla tamamen gevşemez, ancak ton adı verilen bir miktar gerilimi korurlar. Dışa doğru, bu kasların esnekliğinde ifade edilir. İskelet kaslarının tonu, sırayla çeşitli kas liflerini uyaran geniş bir aralıkla birbirini izleyen bireysel sinir uyarılarının kas beslemesi ile ilişkilidir. Bu impulslar, omuriliğin motor nöronlarında ortaya çıkar ve bunların aktivitesi, hem üstteki merkezlerden hem de çevreden, kasların kendisinde bulunan gerilme reseptörlerinden ("kas iğcikleri") gelen impulslarla desteklenir.
İnsanlarda, kas tonusu belirli sınırlar içinde keyfi olarak düzenlenebilir: bir kişi, herhangi bir hareket yapmadan kasları neredeyse tamamen gevşetebilir veya hafifçe zorlayabilir.
Kas çalışması ve gücü. Belirli bir uyarı gücünde kasın kasılma miktarı (kısalma derecesi), hem morfolojik özelliklerine hem de fizyolojik durumuna bağlıdır. Uzun kaslar kısa olanlardan daha fazla kasılır. Kasın orta derecede gerilmesi kasılma etkisini arttırır, kuvvetli gerilme ile kasın kasılması zayıflar. Uzun süreli çalışmanın bir sonucu olarak kas yorgunluğu gelişirse, kasılmasının büyüklüğü düşer.
Kas gücünü ölçmek için kaldırabileceği maksimum yükü belirleyin. Bu kuvvet çok büyük olabilir: Örneğin bir köpek, çene kasları ile vücut ağırlığının 8,3 katı olan bir yükü kaldırabilir. Bir kişinin baldır kaslarının gücü, omuzlarına yerleştirilen ve ayak parmaklarının üzerinde yükselebildiği yükün büyüklüğü ile değerlendirilir.

Pirinç. 6. Çeşitli kasların yapı tipleri (A.A. Ukhtomsky'ye göre).
A - paralel lif akışına sahip kaslar; B - fusiform kas; B - pennat kas
Bir kasın gücü, ceteris paribus, uzunluğuna değil, kesitine bağlıdır: kasın fizyolojik kesiti ne kadar büyükse, yani. tüm liflerinin enine kesitlerinin toplamı, kaldırabileceği yük o kadar büyük olur. Fizyolojik kesit, sadece uzunlamasına düzenlenmiş liflere sahip kaslarda geometrik olanla çakışır; eğik liflere sahip kaslarda, liflerin enine kesitlerinin toplamı, kasın kendisinin geometrik enine kesitini önemli ölçüde aşabilir (Şekil 6). Bu nedenle, eğik liflere sahip bir kasın kuvveti, aynı kalınlıkta fakat boyuna liflere sahip bir kasın kuvvetinden çok daha fazladır. Farklı kasların gücünü karşılaştırmak için, bir kasın kaldırabileceği maksimum yük, mutlak kas gücünü hesaplamak için kesit alanına bölünür. Örneğin, insan gastroknemius kasının mutlak gücü 5,9 kg / cm2, omuz fleksörü - 8.1, çiğneme kası - 10, omuzun pazı - 11.4, omuzun triseps - 16.8, düz kaslar - 1 kg / cm2.
Memelilerin ve insanların kaslarının çoğu tüylü bir yapıya sahiptir. Pennat kas, geniş bir fizyolojik kesite sahiptir ve bu nedenle büyük bir güce sahiptir.
Kasın çalışması, kaldırılan yükün çarpımı ile kasın kısalma miktarı ile ölçülür, yani. kilogram metre veya gram santimetre cinsinden ifade edilir.
Miyoglobinin içeriğine göre kas lifleri kırmızı, beyaz ve orta olarak ayrılır. Kırmızı lifler "yavaş" ve beyaz - "hızlı" olarak kabul edilir. Bazı bilim adamları, insan kaslarının doğumda sadece "yavaş" liflerden oluştuğuna ve bazılarının gelişim sırasında "hızlı" liflere dönüştüğüne inanmaktadır. Diğerleri, kasların yerinin, yapılarının ve işlevlerinin özelliklerinin genetik olarak önceden belirlenmiş olduğuna inanıyor.
Kırmızı lifler esas olarak aerobik modda çalışır, beyaz lifler ise oksijen eksikliği, yani. farklı metabolik süreçler gerçekleştirirler. Kırmızı lifler tipik olarak hafif ila orta dereceli işler için kullanılırken, beyaz lifler ancak çok yoğun çalışma sırasında uyarıcı impulsların akışında önemli bir artıştan sonra işlevsel hale gelir. Ara tip lifler hem kırmızı hem de beyaz liflerin özelliklerini korur ve "hızlı kırmızı" olarak da adlandırılır.
Belirli liflerin yüzdesi, sporcunun uzmanlığını belirler. Kural olarak, ağırlıklı olarak kırmızı kas sahipleri, artan dayanıklılık gerektiren sporlarda (yüzme, bisiklete binme, orta ve uzun mesafelerde koşma vb.) daha iyi sonuçlar elde eder. Daha fazla beyaz kas lifi olanlar, kuvvet antrenmanında daha iyidir. İkincisi, beyaz liflerin daha kolay hipertrofisi (hacimdeki artış) ile de açıklanmaktadır. Ancak, her şey yalnızca doğa tarafından önceden belirlenmiş değildir. Bazı uzmanların kas yapısını şekillendirmede tercih ettiği antrenman faktörleri de vardır.
Kaslar egzersize nasıl alışır? Antrenman sırasında, kaslarda enerji substratlarının depolama ve kullanım mekanizmaları oluşur: kreatin fosfat, glikojen ve yağlar (triaçilgliseritler şeklinde). Kasta çok az ATP ve kreatin fosfat veya fosfajen rezervi vardır. Fosfatlar sürekli olarak sentezlenir (beyaz kas liflerindeki kreatin fosfat içeriği, 1 g ıslak kas kütlesi başına yaklaşık 30 μmol'dür), ancak birkaç saniyeden uzun süren egzersiz sırasında hızla tüketilir. Enerji yoğun glikojen ve triaçilgliseritler, kaslar için rezerv "yakıt" kaynaklarının büyük kısmını oluşturur.
Beyaz kas liflerinde hız ve kuvvet antrenmanının etkisi altında, glikolitik enzimlerin aktivitesi artar ve kaslarda ilgili enzimlerin bulunduğu granüller şeklinde depolanan büyük miktarda glikojen oluşur. Kas hücrelerinin tampon sistemlerinde de değişiklikler vardır: Yoğun çalışma sırasında hücre içinde pH değişir değişmez, bu hemen glikolizden sorumlu enzimlerin çalışmasını etkiler.
Yoğun egzersiz sırasında anaerobik glikolizin sağlayabileceği toplam iş miktarı, glikojen rezervine bağlıdır (moleküllerinin her birinin parçalanması sırasında 6.2 mol ATP oluşur). Kas glikojen depolarının kullanımı, hormonal ve sinirsel uyaranlar tarafından tetiklenir. Bu hormonlardan biri olan adrenalin, ATP yeniden sentezi için glikojen kullanma süreçlerini önemli ölçüde aktive edebilir.
Antrenmanın etkisi altında, kaslardaki anaerobik glikoliz defalarca arttırılabilir. Bu nedenle, bazı raporlara göre, eğitimli sprinterlerde bacak kaslarındaki glikoliz süreçleri iki bin kat artıyor.
Ancak anaerobik glikoliz nedeniyle bir kişi sadece 2-3 dakika egzersiz yapabilir. Bundan sonra, oksidatif fosforilasyon süreçleri kaçınılmaz olarak başlar.
Uzun süreli çalışma sırasında kırmızı ve orta kas lifleri ana karakterler haline gelir. Bu kasların aktivitesi için enerji, yeterli miktarda oksijen varlığında oksidatif enzimlerin yardımıyla mitokondride (kırmızı kas liflerinin hücrelerinde beyazlardan çok daha fazlası vardır) oluşur.
Kısa süreli çalışma sırasında aktif olarak kullanılan glikojen, uzun süreli egzersiz sırasında bile (yağlarla birlikte) ana endojen substrattır. Bu yakıtların her ikisi de, özellikle yağlar, kırmızı ve ara liflerde depolanır. Yağlar, tüketilen oksijen birimi başına enerji çıkışı verimliliğinde glikojenden biraz daha düşüktür: oksitlendiklerinde 5.6 mol ATP oluşur.
Oksidatif fosforilasyon ile kas ayrıca merkezi depolardan (karaciğerden glikojen ve yağ dokusundan yağ) enerji substratları alabilir ve hatta çalışma sırasında dışarıdan gelen enerji kaynaklarını, örneğin bir maraton yarışı sırasında karbonhidrat takviyelerini kullanabilir - doğanın sahip olduğu uzun süreli çalışma için ek fırsatlar sağladı.
Dayanıklılık için antrenman yapan sporcular için uzun süreli ve yoğun kas çalışmasının en iyi enerji ile karbonhidrat ve yağların aynı anda kullanılmasıyla sağlandığı merak edilecektir. Görünüşe göre burada bir paradoks var. Sonuçta, karbonhidratlar sadece 20-30 dakikalık yoğun çalışma için yeterlidir ve yağlar çok daha uzun süre kullanılabilir. Bununla birlikte, mesele şu ki, yağların tek başına kullanılması, aynı anda yağ ve karbonhidrat kullanımına göre yarı yarıya enerji üretim oranı sağlıyor. Ve yapılan işin yoğunluğuna bağlıdır. Bu nedenle, biyokimyacılara göre, aerobik koşullarda yüksek yoğunluklu uzun süreli çalışma sağlamak için glikojen en iyi "yakıt"tır. Bilim adamları, yükün başlamasından önce kaslardaki glikojen içeriğinin tamamen tükenmesine kadar çalışma süresinin bir bağımlılığını bile bulmuşlardır. Ancak yük 2-3 saat devam ederse, vücut kas kasılmalarını sağlamak için hem glikojen hem de yağları kullanmaya başlar. Yağların parçalanmasına geçildiğinde işin gücü azalır. Önce triaçilgliseritler, ardından kandan gelen serbest yağ asitleri kullanılır.
Kuvvet sporları ve vücut geliştiriciler konusunda uzmanlaşmış sporcular, elbette, çalışan kas hipertrofisi konusuyla ilgileniyorlar.
Kasın sistematik yoğun çalışması, kas dokusu kütlesinde bir artışa yol açar. Bu fenomene çalışan kas hipertrofisi denir. Hipertrofi, kas liflerinin protoplazma kütlesindeki bir artışa dayanır ve bu da kalınlaşmalarına neden olur. Bu, proteinlerin ve glikojenin yanı sıra kas kasılması için gerekli enerjiyi sağlayan maddeler - ATP ve kreatin fosfat içeriğini arttırır. Bu nedenle, hipertrofik bir kasın kasılma gücü ve hızı, hipertrofik olmayan bir kastan daha yüksektir.
Eğitimli kişilerde kas dokusu kütlesindeki artış, vücudun kas yapısının vücut ağırlığının% 50'si (normal% 35-40 yerine) olabileceği gerçeğine yol açar.
Bir kişi günlük olarak çok fazla stres (kuvvet yükü) gerektiren uzun süre kas çalışması yaparsa hipertrofi gelişir. Fazla çaba sarf etmeden yapılan kas çalışması, çok uzun sürse bile kas hipertrofisine yol açmaz.
Kas hipertrofisi, kısa süreli "patlayıcı" işler yapmak için çok önemlidir. Belki de bu, insan kaslarındaki kreatin fosfat rezervlerinin, kas kütlesi birimi başına belirli bir miktardan daha fazla artmaması gerçeğinden kaynaklanmaktadır. Bu nedenle, kas hacmindeki bir artış, kaslardaki bu enerji yoğun substratın toplam miktarında bir artışa ve buna bağlı olarak maksimum güç çalışmasını daha verimli bir şekilde gerçekleştirme yeteneğinde bir artışa katkıda bulunur.
Çalışan hipertrofinin tersi, hareketsizlikten kaynaklanan kas atrofisidir. Kasın herhangi bir nedenle normal çalışmasını gerçekleştirme yeteneğini kaybettiği tüm durumlarda, örneğin, bir alçıda bir uzuvun uzun süre hareketsiz kalması, hastanın uzun süre yatakta kalması, bir tendonun kesilmesi ile gelişir. , bunun sonucunda kas yüke karşı iş yapmayı bırakır, vb. .d.
Atrofi ile kas liflerinin çapı ve kasılma proteinlerinin, glikojenin, ATP'nin ve içlerindeki kasılma aktivitesi için önemli olan diğer maddelerin içeriği keskin bir şekilde düşer. Normal kas çalışmasının yeniden başlamasıyla atrofi yavaş yavaş kaybolur.
Kas yorgunluğu.Çalışma veya antrenman sonucunda oluşan ve dinlenme sonrasında kaybolan kas performansında geçici bir azalma yorgunluk ile belirlenir.
Uzun bir süre boyunca küçük bir yükün askıya alındığı izole bir kas ritmik elektriksel uyaranlarla tahriş olursa, kasılmalarının genliği sıfıra ulaşana kadar yavaş yavaş azalır. Bu şekilde elde edilen eğriye yorulma eğrisi denir.
Yorgunluk sırasında kasılmaların genliğinde bir değişiklik ile birlikte, kasılmanın gizli süresi artar ve uyarılabilirlik azalır. Ancak tüm bu değişiklikler, kas çalışmasının başlamasından hemen sonra meydana gelmez - kasılma genliklerinde bir artış ve kas uyarılabilirliğinde hafif bir artış gözlemlenen belirli bir süre vardır. Bu durumda, kas kolayca uzayabilir hale gelir. Bu gibi durumlarda kasın "çalıştığını" söylerler, yani. Belirli bir ritim ve uyarı gücünde çalışmaya uyum sağlar. Daha uzun süreli tahriş ile kas liflerinin yorgunluğu meydana gelir.
Bunun nedeni, kas liflerinin performansı üzerinde iç karartıcı bir etkiye sahip olan kasta metabolik ürünlerin (özellikle glikojenin parçalanması sırasında oluşan laktik asit) birikmesi olabilir. Bu ürünlerin bazıları ve potasyum iyonları, liflerden hücre içi boşluğa yayılır ve uyarılabilir zarın aksiyon potansiyeli oluşturma yeteneği üzerinde iç karartıcı bir etkiye sahiptir.
Ek olarak, kastaki yorgunluğun gelişimi, içindeki enerji rezervlerinin kademeli olarak tükenmesinden etkilenir.
Yukarıdakilerin tümü izole bir kas için geçerlidir. Sonuçta, vücutta çalışırken, kas sürekli olarak kanla beslenir ve bu nedenle belirli miktarda besin (glikoz, amino asitler) alır ve kas liflerinin normal işleyişini bozan metabolik ürünlerden salınır. Temel fark, vücutta uyarıcı dürtülerin sinirden kasa gelmesidir. Nöromüsküler kavşak, kas liflerinden çok daha erken yorulur ve bu nedenle sinirden kasa uyarı iletimini bloke ederek, kas liflerini uzun süreli çalışmanın neden olduğu yorgunluktan korur. Tüm organizmada, nöromüsküler bağlantılardan önce bile sinir merkezleri çalışma sırasında yorulur.
Yük kaldırmak için uzun bir çalışmadan sonra insan elinin yorgun kaslarının çalışma kapasitesinin geri kazanılması, dinlenme sırasında diğer el veya alt uzuvlarla iş yapılırsa hızlandırılabilir.
Devam edecek
İç organlar, deri, kan damarları.
İskelet kaslarıİskelet ile birlikte, vücudun duruşunu koruyan ve vücudu uzayda hareket ettiren kas-iskelet sistemini oluştururlar. Ek olarak, iç organları hasardan koruyan koruyucu bir işlev görürler.
İskelet kasları, kemikleri ve eklemlerini, bağlarını ve tendonlarını da içeren kas-iskelet sisteminin aktif kısmıdır. Kas kütlesi toplam vücut ağırlığının %50'sine ulaşabilir.
Fonksiyonel bir bakış açısından, kas liflerine sinir uyarıları gönderen motor nöronlar da motor aparata atfedilebilir. İskelet kaslarını aksonlarla innerve eden motor nöronların gövdeleri omuriliğin ön boynuzlarında, maksillofasiyal bölgenin kaslarını innerve edenler ise beyin sapının motor çekirdeklerinde bulunur. Motor nöron aksonu iskelet kasının girişinde dallanır ve her dal ayrı bir kas lifi üzerinde nöromüsküler sinaps oluşumunda rol oynar (Şekil 1).
Pirinç. 1. Bir motor nöronun aksonunun akson terminallerine dallanması. elektronogram

Pirinç. İnsan iskelet kasının yapısı
İskelet kasları, kas demetleri halinde birleştirilen kas liflerinden oluşur. Bir motor nöronun akson dalları tarafından innerve edilen kas lifleri grubuna motor (veya motor) ünite denir. Göz kaslarında 1 motor ünite 3-5 kas lifi, gövde kaslarında - yüzlerce lif, soleus kasında - 1500-2500 lif içerebilir. 1. motor ünitenin kas lifleri aynı morfofonksiyonel özelliklere sahiptir.
iskelet kası fonksiyonlarışunlardır:
- vücudun uzayda hareketi;
- akciğerlerin havalandırılmasını sağlayan solunum hareketlerinin uygulanması da dahil olmak üzere vücut bölümlerinin birbirine göre hareketi;
- vücut pozisyonunu ve duruşunu korumak.
İskelet kasları, iskelet ile birlikte vücudun duruşunu koruyan ve vücudu uzayda hareket ettiren kas-iskelet sistemini oluşturur. Bununla birlikte, iskelet kasları ve iskelet, iç organları hasardan koruyan koruyucu bir işlev görür.
Ek olarak, çizgili kaslar, sıcaklık homeostazını korumak için ısı üretmede ve belirli besin maddelerini depolamada önemlidir.

Pirinç. 2. İskelet kası fonksiyonları
İskelet kaslarının fizyolojik özellikleri
İskelet kasları aşağıdaki fizyolojik özelliklere sahiptir.
uyarılabilirlik. Plazma zarının (sarkolemma) bir sinir impulsunun gelişine uyarma ile yanıt verme özelliği ile sağlanır. Çizgili kas liflerinin (E 0 yaklaşık 90 mV) zarının dinlenme potansiyelindeki daha büyük fark nedeniyle, uyarılabilirlikleri sinir liflerininkinden (E 0 yaklaşık 70 mV) daha düşüktür. Aksiyon potansiyeli genliği, diğer uyarılabilir hücrelerden daha büyüktür (yaklaşık 120 mV).
Bu, iskelet farelerinin biyoelektrik aktivitesini kaydetmeyi pratikte oldukça kolaylaştırır. Aksiyon potansiyelinin süresi, kas liflerinin uyarılmış zarının mutlak refrakterlik fazının kısa süresini belirleyen 3-5 ms'dir.
İletkenlik. Plazma zarının yerel dairesel akımlar oluşturması, bir aksiyon potansiyeli oluşturması ve yürütmesi özelliği ile sağlanır. Sonuç olarak, aksiyon potansiyeli zar boyunca kas lifi boyunca ve zarın oluşturduğu enine tübüllerin derinliklerinde yayılır. Aksiyon potansiyelinin hızı 3-5 m/s'dir.
kasılma. Membranın uyarılmasının ardından uzunluklarını ve gerilimlerini değiştirmek, kas liflerinin spesifik bir özelliğidir. Kasılma, kas lifinin özel kasılma proteinleri tarafından sağlanır.
İskelet kasları da kas gevşemesi için önemli olan viskoelastik özelliklere sahiptir.

Pirinç. İnsan iskelet kasları
İskelet kaslarının fiziksel özellikleri
İskelet kasları, uzayabilirlik, esneklik, güç ve iş yapabilme yeteneği ile karakterize edilir.
genişletilebilirlik - bir kasın bir çekme kuvvetinin etkisi altında uzunluğunu değiştirme yeteneği.
esneklik - Bir kasın, bir gerilme veya deforme edici kuvvetin kesilmesinden sonra orijinal şeklini geri kazanma yeteneği.
- bir kasın bir yükü kaldırma yeteneği. Farklı kasların güçlerini karşılaştırmak için, maksimum kütlenin fizyolojik kesitinin santimetre kare sayısına bölünmesiyle spesifik güçleri belirlenir. İskelet kasının gücü birçok faktöre bağlıdır. Örneğin, belirli bir zamanda uyarılan motor ünitelerinin sayısı. Aynı zamanda motor ünitelerinin senkronizasyonuna da bağlıdır. Kasın gücü ayrıca başlangıç uzunluğuna da bağlıdır. Kasın maksimum kasılma geliştirdiği belirli bir ortalama uzunluk vardır.
Düz kasların gücü ayrıca başlangıç uzunluğuna, kas kompleksinin uyarılma senkronizasyonuna ve ayrıca hücre içindeki kalsiyum iyonlarının konsantrasyonuna da bağlıdır.
kas yeteneği işi yapmak. Kasın çalışması, kaldırılan yükün kütlesi ile kaldırma yüksekliğinin çarpımı ile belirlenir.
Kaldırılan yükün kütlesindeki bir artışla kas çalışması artar, ancak belirli bir sınıra kadar, daha sonra yükteki bir artış işte bir azalmaya yol açar, yani. kaldırma yüksekliği azalır. Maksimum iş, orta yüklerde kas tarafından yapılır. Buna ortalama yükler yasası denir. Kas çalışmasının miktarı, kas liflerinin sayısına bağlıdır. Kas ne kadar kalınsa o kadar fazla ağırlık kaldırabilir. Uzun süreli kas gerginliği yorgunluğa yol açar. Bunun nedeni kastaki (ATP, glikojen, glikoz) enerji rezervlerinin tükenmesi, laktik asit ve diğer metabolitlerin birikmesidir.
İskelet kaslarının yardımcı özellikleri
Uzatılabilirlik, bir kasın bir çekme kuvvetinin etkisi altında uzunluğunu değiştirme yeteneğidir. Elastikiyet - bir kasın bir gerilme veya deforme edici kuvvetin kesilmesinden sonra orijinal uzunluğunu alma yeteneği. Canlı bir kas küçük ama mükemmel bir esnekliğe sahiptir: küçük bir kuvvet bile kasın nispeten büyük bir uzamasına neden olabilir ve orijinal boyutlarına dönüşü tamamlanmıştır. Bu özellik, iskelet kaslarının normal fonksiyonları için çok önemlidir.
Bir kasın gücü, kasın kaldırabileceği maksimum yük ile belirlenir. Farklı kasların güçlerini karşılaştırmak için özel güçleri belirlenir, yani. kasın kaldırabileceği maksimum yük, fizyolojik kesitinin santimetre kare sayısına bölünür.
Bir kasın iş yapabilme yeteneği. Kasın çalışması, kaldırılan yükün değerinin kaldırma yüksekliği ile çarpımı ile belirlenir. Kasın çalışması, yükteki bir artışla kademeli olarak artar, ancak belirli bir sınıra kadar, bundan sonra yükteki bir artış, yükün yüksekliği azaldığı için işin azalmasına neden olur. Sonuç olarak, kasın maksimum çalışması orta yüklerde gerçekleştirilir.
Kas yorgunluğu. Kaslar sürekli çalışamaz. Uzun süreli çalışma performanslarında düşüşe yol açar. Uzun süreli çalışma sırasında oluşan ve istirahatten sonra kaybolan kas performansındaki geçici azalmaya kas yorgunluğu denir. İki tür kas yorgunluğu arasında ayrım yapmak gelenekseldir: yanlış ve doğru. Sahte yorgunlukta yorulan kas değil, sinaps adı verilen uyarıları sinirden kasa iletmek için özel bir mekanizmadır. Sinapsta, nörotransmiterlerin rezervleri tükenir. Gerçek yorgunluk ile kasta aşağıdaki süreçler meydana gelir: yetersiz oksijen kaynağı nedeniyle besinlerin az oksitlenmiş parçalanma ürünlerinin birikmesi, kas kasılması için gerekli enerji kaynaklarının tükenmesi. Yorgunluk, kas kasılmasının gücünde ve kas gevşeme derecesinde bir azalma ile kendini gösterir. Kas bir süre çalışmayı durdurur ve dinlenirse, sinaps çalışması geri yüklenir ve kanla birlikte metabolik ürünler çıkarılır ve besinler verilir. Böylece kas kasılma ve iş üretme yeteneğini yeniden kazanır.
Tek kesim
Tek bir uyaranla innerve eden bir kas veya motor sinirin tahrişi, tek bir kas kasılmasına neden olur. Böyle bir kasılmanın üç ana aşaması vardır: latent faz, kısalma fazı ve gevşeme fazı.
İzole edilmiş bir kas lifinin tek bir kasılmasının genliği, stimülasyonun gücüne bağlı değildir, yani. ya hep ya hiç yasasına uyar. Bununla birlikte, birçok liften oluşan tüm kasın doğrudan tahrişi ile kasılması, tahrişin gücüne bağlıdır. Eşik akım gücünde, reaksiyona sadece az sayıda lif katılır, bu nedenle kas kasılması zar zor fark edilir. Stimülasyonun gücündeki bir artışla, uyarma tarafından kapsanan liflerin sayısı artar; tüm lifler kasılana kadar kasılma artar ("maksimum kasılma") - bu etkiye Bowditch merdiveni denir. Tahriş edici akımın daha fazla amplifikasyonu kas kasılmasını etkilemez.

Pirinç. 3. Tek bir kas kasılması: A - kas tahrişi anı; a-6 - gizli dönem; 6-in - azaltma (kısaltma); cd - gevşeme; d-e - ardışık elastik salınımlar.
tetanoz kasları
Doğal koşullar altında, merkezi sinir sisteminden gelen iskelet kası, kendisi için yeterli uyaran görevi gören tek uyarma dürtülerini değil, kasın uzun süreli bir kasılma ile yanıt verdiği bir dizi uyarıyı alır. Ritmik uyarıya tepki olarak ortaya çıkan kasın uzun süreli kasılmasına tetanik kasılma veya tetanoz denir. İki tür tetanoz vardır: tırtıklı ve pürüzsüz (Şekil 4).
pürüzsüz tetanoz her bir sonraki uyarma darbesi kısalma aşamasına girdiğinde meydana gelir ve pürüzlü - gevşeme aşamasında.
Tetanik kasılmanın genliği, tek bir kasılmanın genliğini aşıyor. Akademisyen N.E. Vvedensky, tetanoz genliğinin değişkenliğini kas uyarılabilirliğinin eşit olmayan değeriyle doğruladı ve fizyolojiye uyarılma sıklığında optimum ve kötümser kavramlarını tanıttı.
En uygun her bir sonraki tahrişin artan kas uyarılabilirliği aşamasına girdiği böyle bir tahriş sıklığı olarak adlandırılır. Aynı zamanda, maksimum boyutta (optimal) bir tetanoz gelişir.
kötümser her bir sonraki tahrişin kasın azaltılmış uyarılabilirliğinin bir aşamasında gerçekleştirildiği böyle bir tahriş sıklığı olarak adlandırılır. Bu durumda tetanoz değeri minimum (kötümser) olacaktır.

Pirinç. 4. Farklı uyarı frekanslarında iskelet kasının kasılması: I - kasın kasılması; II - tahriş sıklığını işaretleyin; a - tek kasılmalar; b- dentat tetanoz; c - pürüzsüz tetanoz
Kas kasılma modları
İskelet kasları izotonik, izometrik ve karışık kasılma modları ile karakterize edilir.
saat izotonik kasın kasılması uzunluğunu değiştirir ve gerilim sabit kalır. Böyle bir kasılma, kas direncin üstesinden gelmediğinde (örneğin, yükü hareket ettirmediğinde) meydana gelir. Doğal koşullarda izotonik tipe yakın kasılmalar dil kaslarının kasılmalarıdır.
saat eş ölçülü aktivitesi sırasında kasta kasılma, gerginlik artar, ancak kasın her iki ucunun sabitlenmesi nedeniyle (örneğin, kas büyük bir yükü kaldırmaya çalışıyor), kısalmaz. Kas liflerinin uzunluğu sabit kalır, sadece gerginliklerinin derecesi değişir.
Benzer mekanizmalarla indirgenirler.
Vücutta kas kasılmaları asla tamamen izotonik veya izometrik değildir. Her zaman karışık bir karaktere sahiptirler, yani. kasın hem uzunluğunda hem de gerginliğinde eşzamanlı bir değişiklik vardır. Bu azaltma modu denir oksotonik, kas gerginliği baskınsa veya oksometrik, kısalma geçerliyse.
Aynı metalin birkaç biyokimyasal görevi yerine getirdiğini defalarca fark etme fırsatı bulduk: demir oksijen ve elektronları taşır, bakır benzer işlemlere katılır, çinko polipeptitlerin hidrolizini ve bikarbonatların ayrışmasını destekler, vb.
Ancak kalsiyum bu konuda tüm rekorları kırıyor. Kalsiyum iyonları, birikimleri muazzam boyutlara ulaşan mercanlarda koruyucu kabuklar oluşturur; kas aktivitesi sağlayan enzimlerin çalışması için kalsiyum gereklidir; kalsiyum kan pıhtılaşma sistemini düzenler, bazı enzimleri aktive eder; aynı zamanda omurgalıların vb. kemiklerinin ve dişlerinin bir parçasıdır.
Kalsiyum döngüsü, karbonat tuzlarının farklı çözünürlüğü ile kolaylaştırılır: CaCO 3 karbonat suda az çözünür ve Ca(HCO 3) 2 bikarbonat oldukça çözünür ve çözeltideki konsantrasyonu karbondioksit konsantrasyonuna bağlıdır ve bu nedenle , bu gazın çözelti üzerindeki kısmi basıncı üzerine ; bu nedenle, dağ kaynaklarının karbonik suları yeryüzüne aktığında ve karbondioksiti (karbondioksit) kaybettiğinde, kalsiyum karbonat çökerek kristal kümeler (mağaralarda sarkıt ve dikitler) oluşturur. Mikroorganizmalar, deniz suyundan bikarbonat çıkararak ve koruyucu kabuklar oluşturmak için karbonat kullanarak benzer bir işlem gerçekleştirir.
Daha yüksek hayvanların organizmalarında kalsiyum, mekanik olarak güçlü yapıların yaratılmasıyla ilgili işlevleri de yerine getirir. Kemiklerde kalsiyum, bileşimde mineral apatit 3Ca 3 (PO 4) 2 * CaF 2 (Cl) ile benzer şekilde tuzlar şeklinde bulunur. Parantez içindeki klor sembolü, bu mineralde flor yerine klorun kısmi ikamesini gösterir.
Kemik dokusunun oluşumu, D grubu vitaminlerinin etkisi altında gerçekleşir; bu vitaminler, sırayla, güneşten gelen ultraviyole radyasyonun etkisi altındaki organizmalarda sentezlenir. Balık yağında önemli miktarda D vitamini bulunur, bu nedenle bebek mamasındaki vitamin eksikliği ile kalsiyum bağırsaklarda emilmez ve raşitizm semptomları gelişir; doktorlar bir ilaç olarak balık yağı veya saf D vitamini müstahzarları reçete eder.Bu vitaminin fazlalığı çok tehlikelidir: tersine işleme neden olabilir - kemik dokusunun çözünmesi!
Gıda ürünlerinden kalsiyum, sütte, süt ürünlerinde (özellikle süt proteini kazeini kalsiyum iyonları ile ilişkili olduğundan süzme peynirde) ve bitkilerde bulunur.
Küçük bir moleküler ağırlığa (yaklaşık 11.000) sahip olan ve balıkların kaslarında bulunan proteinler, kalsiyum iyonlarını aktif olarak yakalama kabiliyeti gösterir. Bazıları (örneğin, sazan albümini) kapsamlı bir şekilde incelenmiştir; bileşimlerinin olağandışı olduğu ortaya çıktı: alanin ve fenilalanin amino asitlerinin çoğunu içerirler ve histidin, sistein ve arginin içermezler - diğer proteinlerin neredeyse değişmemiş bileşenleri.
Kalsiyum iyonunun karmaşık bileşikleri için, köprü oluşumu karakteristiktir - iyon, elde edilen komplekste esas olarak karboksil ve karbonil gruplarını bağlar.
Kalsiyum iyonunun koordinasyon sayısı büyüktür ve sekize ulaşır. Görünüşe göre, bu özelliği, vücut için önemli olan nükleik asitlerin (RNA) hidroliz sürecini katalize eden enzim ribonükleazın etkisinin altında ve enerji salınımının altında yatmaktadır. Kalsiyum iyonunun bir su molekülü ile bir fosfat grubunu bir araya getiren katı bir kompleks oluşturduğu varsayılır; bir kalsiyum iyonu ile çevrili arginin kalıntıları, fosfat grubunun sabitlenmesine katkıda bulunur. Kalsiyum tarafından polarize edilir ve su molekülü tarafından daha kolay saldırıya uğrar. Sonuç olarak, fosfat grubu nükleotitten ayrılır. Bu enzimatik reaksiyondaki kalsiyum iyonunun aynı oksidasyon durumundaki diğer iyonlarla değiştirilemeyeceği de kanıtlanmıştır.
Kalsiyum iyonları ayrıca diğer enzimleri, özellikle a-amilazı (nişastanın hidrolizini katalize eder) aktive eder, ancak bu durumda, kalsiyum yapay koşullar altında üç yüklü neodimyum metal iyonu ile yine de değiştirilebilir.
Kalsiyum aynı zamanda en çok bir makineye benzeyen bu şaşırtıcı biyolojik sistemin en önemli bileşenidir - kas sistemi. Bu makine, gıda maddelerinin içerdiği kimyasal enerjiden mekanik iş üretir; verimliliği yüksektir; neredeyse anında bir dinlenme durumundan hareket durumuna aktarılabilir (ayrıca, dinlenme durumunda hiçbir enerji tüketilmez); özgül gücü 1 kg kütle başına yaklaşık 1 kW'dır, hareketlerin hızı iyi düzenlenmiştir; makine tekrarlayan hareketler gerektiren uzun süreli işler için oldukça uygundur, hizmet ömrü yaklaşık 2.6*106 işlemdir. Yaklaşık olarak kas prof tanımladı. Wilkie, popüler bir derste, bir makinenin ("doğrusal motor") yiyecek olarak hizmet edebileceğini ekleyerek.
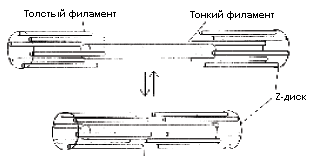
Bilim adamlarının bu "doğrusal motorun" içinde ne olduğunu, bir kimyasal reaksiyonun nasıl amaca yönelik hareketi oluşturduğunu ve tüm bunlarda kalsiyum iyonlarının nasıl bir rol oynadığını anlaması çok zordu. Kas dokusunun bir zar (sarkolemma) ile çevrili liflerden (uzun hücreler) oluştuğu artık tespit edilmiştir. Kas hücrelerinde miyofibriller vardır - kasın sıvıya batırılmış kasılma elemanları - sarkoplazma. Miyofibriller sarkomer adı verilen bölümlerden oluşur. Sarkomerler iki tip filament sistemi içerir - kalın ve ince.
Kalın filamentler, protein miyozinden oluşur. Miyozin molekülleri, bir ucunda kalınlaşma olan uzun parçacıklardır - bir kafa. Başlar, ipliksi molekülün yüzeyinin üzerinde çıkıntı yapar ve molekül eksenine farklı açılarda yerleştirilebilir. Miyozinin moleküler ağırlığı 470.000'dir.
İnce filamentler, küresel bir şekle sahip aktin protein molekülleri tarafından oluşturulur. Aktinin moleküler ağırlığı 46.000'dir.Aktin partikülleri, uzun bir çift sarmal elde edilecek şekilde düzenlenmiştir. Her yedi aktin molekülü, başka bir proteinin, troponin'in küresel bir molekülünü taşıyan (uçlardan birine daha yakın) tropomiyosin proteininin filamentli bir molekülü ile bağlanır (Şekil 19). İnce bir iskelet kası filamenti, 400'e kadar aktin molekülü ve 60'a kadar tropomiyozin molekülü içerir. Böylece kasın çalışması, dört proteinden oluşan parçaların etkileşimine dayanır.
İpliklerin eksenlerine dik olan protein oluşumları - bir ucuna ince ipliklerin tutturulduğu z-plakaları. İnce ipler arasına kalın ipler yerleştirilir. Gevşemiş bir kasta z-plakaları arasındaki mesafe yaklaşık 2,2 mikrondur. Kas kasılması, bir sinir impulsunun etkisi altında, miyozin moleküllerinin çıkıntılarının (başlarının) ince filamentlere bağlanması ve sözde çapraz bağlar veya köprülerin ortaya çıkmasıyla başlar. Plakanın her iki tarafındaki kalın filamentlerin başları zıt yönlerde eğimlidir, bu nedenle dönerek kalın olanlar arasında ince bir iplik çekerler ve bu da tüm kas lifinin kasılmasına neden olur.
Kas çalışması için enerji kaynağı, adenozin trifosforik asidin (ATP) hidroliz reaksiyonudur; bu maddenin varlığı kas sisteminin çalışması için gereklidir.
1939'da V. A. Engelgardt ve M. N. Lyubimova, miyosin ve aktin - aktomiyosin ile kompleksinin, genel olarak hidrolitik reaksiyonları kolaylaştıran magnezyumun yanı sıra kalsiyum ve potasyum iyonlarının varlığında ATP'nin hidrolizini hızlandıran katalizörler olduğunu kanıtladı. Kalsiyumun özel rolü, aktin ve miyozin arasında çapraz bağların (köprüler) oluşumunu düzenlemesidir. ATP molekülü, kalın filamentlerde miyozin molekülünün başına bağlanır. Daha sonra, bu kompleksi aktif, ancak kararsız bir duruma getiren bir tür kimyasal değişim meydana gelir. Böyle bir kompleks, bir aktin molekülü ile (ince bir iplik üzerinde) temas ederse, ATP hidroliz reaksiyonu nedeniyle enerji açığa çıkacaktır. Bu enerji, köprünün sapmasına ve kalın ipliği protein plakasına yaklaştırmasına, yani kas lifinin kasılmasına neden olur. Daha sonra, yeni bir ATP molekülü aktin-miyozin kompleksine katılır ve kompleks hemen parçalanır: aktin miyozinden ayrılır, köprü artık kalın ipliği ince olana bağlamaz - kas gevşer ve miyozin ve ATP bir zincire bağlı kalır. etkin olmayan bir durumda olan kompleks.
Kalsiyum iyonları, tek bir kas lifini çevreleyen tübüllerde ve veziküllerde bulunur. İnce zarlardan oluşan bu tüp ve vezikül sistemine sarkoplazmik retikulum denir; ipliklerin bulunduğu sıvı bir ortama daldırılır. Bir sinir impulsunun etkisi altında, zarların geçirgenliği değişir ve sarkoplazmik retikulumdan ayrılan kalsiyum iyonları çevreleyen sıvıya girer. Kalsiyum iyonlarının troponin ile birleştiğinde, filamentli tropomiyozin molekülünün konumunu etkilediği ve onu aktif ATP-miyosin kompleksinin aktine bağlanabileceği bir konuma aktardığı varsayılmaktadır. Görünüşe göre, kalsiyum iyonlarının düzenleyici etkisi, tropomiyozin filamentleri yoluyla aynı anda yedi aktin molekülüne kadar uzanır.
Kas kasılmasından sonra, kalsiyum çok hızlı bir şekilde (saniyenin kesirleri) sıvıdan çıkarılır, yine sarkoplazmik retikulumun veziküllerine bırakılır ve kas lifleri gevşer. Sonuç olarak, "doğrusal motorun" çalışma mekanizması, kalın miyozin filamentlerinden oluşan bir sistemin protein plakalarına bağlı ince aktin filamentleri arasındaki boşluğa dönüşümlü olarak itilmesinden oluşur ve bu işlem, sarkoplazmik retikulumdan periyodik olarak çıkan kalsiyum iyonları tarafından düzenlenir ve tekrar bırakıyorum.
İçeriği kastaki kalsiyum içeriğinden çok daha büyük olan potasyum iyonları, küresel aktin formunun filamentli - fibriller forma dönüşmesine katkıda bulunur: bu durumda aktin, miyozin ile daha kolay etkileşime girer.
Bu açıdan bakıldığında, potasyum iyonlarının neden kalp kasının kasılmasını arttırdığı, genel olarak vücudun kas sisteminin gelişimi için neden gerekli olduğu ortaya çıkıyor.
Kalsiyum iyonları, kan pıhtılaşma sürecinde aktif katılımcılardır. Bu sürecin organizmanın yaşamının korunması için ne kadar önemli olduğunu söylemeye gerek yok. Kan pıhtılaşma yeteneğinden yoksun olsaydı, küçük bir çizik yaşam için ciddi bir tehdit oluştururdu. Ancak normal bir vücutta küçük yaralardan kanama 3-4 dakika sonra durur. Hasarlı dokularda yoğun bir fibrin proteini pıhtısı oluşur ve yarayı tıkar. Bir kan pıhtısı oluşumu üzerine yapılan bir araştırma, çeşitli proteinler ve özel enzimler de dahil olmak üzere, oluşumunda karmaşık sistemlerin yer aldığını göstermiştir. Tüm sürecin doğru ilerlemesi için en az 13 faktörün birlikte hareket etmesi gerekir.
Dolaşım sisteminin bir damarı hasar gördüğünde, tromboplastin proteini kan dolaşımına girer. Kalsiyum iyonları, bu proteinin protrombin (yani "trombin kaynağı") adı verilen bir madde üzerindeki etkisinde yer alır. Başka bir protein (globulin sınıfından) protrombinin trombine dönüşümünü hızlandırır. Trombin, molekülleri filamentli bir yapıya sahip olan yüksek moleküler ağırlıklı bir protein olan (molekül ağırlığı yaklaşık 400.000) fibrinojen üzerinde etki eder. Fibrinojen karaciğerde üretilir ve çözünür bir proteindir. Bununla birlikte, trombinin etkisi altında önce monomerik bir forma dönüşür ve sonra polimerize olur ve çözünmeyen bir fibrin formu elde edilir - kanamayı durduran aynı pıhtı. Çözünmeyen fibrin oluşumu sürecinde yine kalsiyum iyonları katılır.
Hareketlilik, tüm yaşam formlarının karakteristik bir özelliğidir. Yönlendirilmiş hareket, hücre bölünmesi, moleküllerin aktif taşınması, protein sentezi sırasında ribozomların hareketi, kas kasılması ve gevşemesi sırasında kromozomların ayrılmasıyla oluşur. Kas kasılması biyolojik hareketliliğin en gelişmiş şeklidir. Kas hareketi de dahil olmak üzere herhangi bir hareket, ortak moleküler mekanizmalara dayanır.
İnsanlarda birkaç çeşit kas dokusu vardır. Çizgili kas dokusu iskelet kaslarını (isteyerek kasabileceğimiz iskelet kası) oluşturur. Düz kas dokusu, iç organların kaslarının bir parçasıdır: gastrointestinal sistem, bronşlar, idrar yolu, kan damarları. Bu kaslar, bilincimizden bağımsız olarak istemsiz olarak kasılır.
Bu derste, spor biyokimyası için en büyük ilgi alanı olan iskelet kaslarının kasılma ve gevşeme yapı ve süreçlerini ele alacağız.
mekanizma kas kasılması bugüne kadar tam olarak açıklanmadı.
Aşağıdakiler iyi bilinmektedir.
1. ATP molekülleri, kas kasılması için enerji kaynağıdır.
2. ATP hidrolizi, enzimatik aktiviteye sahip olan miyozin tarafından kas kasılması sırasında katalize edilir.
3. Kas kasılması için tetikleyici mekanizma, sinir motor dürtüsünün neden olduğu miyositlerin sarkoplazmasındaki kalsiyum iyonlarının konsantrasyonundaki bir artıştır.
4. Kas kasılması sırasında, miyofibrillerin ince ve kalın filamentleri arasında çapraz köprüler veya yapışıklıklar ortaya çıkar.
5. Kas kasılması sırasında, ince iplikler kalın iplikler boyunca kayar, bu da miyofibrillerin ve bir bütün olarak tüm kas lifinin kısalmasına neden olur.
Kas kasılma mekanizmasını açıklayan birçok hipotez vardır, ancak en makul olanı sözde olanıdır. "kayan iplikler" veya "kürek hipotezi" hipotezi (teorisi).
Dinlenme kasında ince ve kalın filamentler birbirinden kopuk durumdadır.
Sinir impulsunun etkisi altında, kalsiyum iyonları sarkoplazmik retikulumun sarnıçlarından ayrılır ve ince filamentlerin proteinine - troponin bağlanır. Bu protein konfigürasyonunu değiştirir ve aktin konfigürasyonunu değiştirir. Sonuç olarak, ince filamentlerin aktinleri ile kalın filamentlerin miyozini arasında enine bir köprü oluşur. Bu, miyozinin ATPaz aktivitesini arttırır. Miyozin ATP'yi parçalar ve bu durumda açığa çıkan enerji nedeniyle miyozin başı bir menteşe veya kayık kürek gibi dönerek kas filamentlerinin birbirine doğru kaymasına neden olur.
Bir dönüş yaptıktan sonra, iplikler arasındaki köprüler kırılır. Miyozinin ATPaz aktivitesi keskin bir şekilde azalır ve ATP hidrolizi durur. Bununla birlikte, sinir impulsunun daha fazla gelmesiyle, yukarıda açıklanan işlem tekrarlandığından, enine köprüler tekrar oluşur.
Her kasılma döngüsünde 1 molekül ATP tüketilir.
Kas kasılması iki işleme dayanır:
kasılma proteinlerinin sarmal bükülmesi;
miyozin zinciri ve aktin arasındaki kompleksin döngüsel olarak tekrar eden oluşumu ve ayrışması.
Kas kasılması, işlevi impulsları iletmek olan nörohormon asetilkolinin salındığı motor sinirin uç plakasına bir aksiyon potansiyelinin gelmesiyle başlatılır. İlk olarak, asetilkolin, sarkolemma boyunca bir aksiyon potansiyelinin yayılmasına yol açan asetilkolin reseptörleri ile etkileşime girer. Bütün bunlar, sarkolemmanın iç yüzeyindeki negatif yükü nötralize ederek, kas lifine akan Na + katyonları için sarkolemmanın geçirgenliğinde bir artışa neden olur. Sarkoplazmik retikulumun enine tübülleri, uyarma dalgasının yayıldığı sarkolemmaya bağlanır. Tübüllerden, uyarma dalgası, aktin ve miyozin filamentlerinin etkileşiminin meydana geldiği alanlarda miyofibrilleri ören veziküllerin ve sarnıçların zarlarına iletilir. Sarkoplazmik retikulumun sarnıçlarına bir sinyal iletildiğinde, ikincisi içlerinde bulunan Ca2+'yı serbest bırakmaya başlar. Salınan Ca2+, tropomiyosine ve daha sonra aktine iletilen konformasyonel kaymalara neden olan Tn-C'ye bağlanır. Aktin, olduğu gibi, içinde bulunduğu ince filamentlerin bileşenleri ile kompleksten salınır. Daha sonra, aktin miyozin ile etkileşime girer ve bu etkileşimin sonucu, ince filamentlerin kalın olanlar boyunca hareket etmesini sağlayan adezyonların oluşumudur.
Kuvvet oluşumu (kısalma), miyozin ve aktin arasındaki etkileşimin doğası gereğidir. Miyozin çubuğu, miyozinin küresel başı belirli bir aktin alanına bağlandığında dönmenin meydana geldiği bölgede hareketli bir menteşeye sahiptir. Aktin filamentlerinin (ince filamentler) H bölgesine geri çekilmesinin nedeni, miyozin ve aktin arasındaki birçok etkileşim bölgesinde aynı anda meydana gelen bu rotasyonlardır. Burada şekilde gösterildiği gibi (maksimum kısalmada) temas ederler veya hatta birbirleriyle örtüşürler.


 içinde
içinde
Resim. Azaltma mekanizması: a- dinlenme durumu; b- orta derecede kasılma; içinde- maksimum kasılma
Bu işlem için enerji, ATP'nin hidrolizi ile sağlanır. ATP, miyozin ATPaz'ın aktif merkezinin bulunduğu miyozin molekülünün başına bağlandığında, ince ve kalın filamentler arasında bağlantı oluşmaz. Görünen kalsiyum katyonu, ATP'nin negatif yükünü nötralize ederek, miyozin ATPaz'ın aktif merkezi ile yakınsamayı teşvik eder. Sonuç olarak, miyozinin fosforilasyonu meydana gelir, yani miyozin, aktin ile adezyonlar oluşturmak ve ince bir filamenti hareket ettirmek için kullanılan enerji ile yüklenir. İnce iplik bir "adım" ilerledikten sonra, ADP ve fosforik asit aktomiyosin kompleksinden ayrılır. Daha sonra miyozin başına yeni bir ATP molekülü eklenir ve tüm süreç miyozin molekülünün bir sonraki başı ile tekrarlanır.
ATP tüketimi de kas gevşemesi için gereklidir. Motor impuls eyleminin sona ermesinden sonra, Ca2+ sarkoplazmik retikulumun sarnıçlarına geçer. Th-C ilişkili kalsiyumu kaybederek troponin-tropomiyozin kompleksinde konformasyonel kaymalara neden olur ve Th-I tekrar aktin aktif bölgelerini kapatarak onları miyozin ile etkileşime giremez hale getirir. Kasılma proteinleri bölgesindeki Ca2+ konsantrasyonu eşiğin altına düşer ve kas lifleri aktomiyosin oluşturma yeteneklerini kaybeder.
Bu koşullar altında, kasılma anında deforme olan stromanın elastik kuvvetleri devreye girer ve kas gevşer. Bu durumda, A diskinin kalın dişleri, H bölgesi ve I diski arasındaki boşluktan ince ipler çıkarılır, orijinal uzunluklarını alır, Z çizgileri birbirinden aynı mesafede uzaklaşır. Kas incelir ve uzar.
hidroliz oranı ATP kas çalışması sırasında çok büyük: 1 dakikada 1 g kas başına 10 mikromole kadar. Genel hisse senetleri ATP Bu nedenle, kasların normal işleyişini sağlamak için küçüktür ATP tüketildiği oranda geri yüklenmelidir.
Kas gevşemesi uzun bir sinir impulsunun alınmasının kesilmesinden sonra ortaya çıkar. Aynı zamanda, sarkoplazmik retikulum sarnıçlarının duvarının geçirgenliği azalır ve kalsiyum pompasının etkisi altındaki kalsiyum iyonları, ATP'nin enerjisini kullanarak sarnıçlara girer. Motor impulsun kesilmesinden sonra kalsiyum iyonlarının retikulum sarnıçlarına çıkarılması önemli bir enerji harcaması gerektirir. Kalsiyum iyonlarının uzaklaştırılması daha yüksek bir konsantrasyon yönünde gerçekleştiğinden, yani. ozmotik gradyana karşı, daha sonra her bir kalsiyum iyonunu çıkarmak için iki ATP molekülü harcanır. Sarkoplazmadaki kalsiyum iyonlarının konsantrasyonu hızla başlangıç seviyesine düşer. Proteinler, dinlenme durumunun konformasyon özelliğini yeniden kazanır.






 Athletico Futbol Kulübü
Athletico Futbol Kulübü Ksenia Sheveleva: “Çalışana kadar hayaller işe yaramaz!
Ksenia Sheveleva: “Çalışana kadar hayaller işe yaramaz! Resmi spor müsabakalarının yapıldığı günlerde ziyaret etme yasağı şeklinde idari ceza verme prosedürü
Resmi spor müsabakalarının yapıldığı günlerde ziyaret etme yasağı şeklinde idari ceza verme prosedürü Aşk, havuç: ünlü fitness çiftleri Kseniya sheveleva fitness bikini yaşı
Aşk, havuç: ünlü fitness çiftleri Kseniya sheveleva fitness bikini yaşı